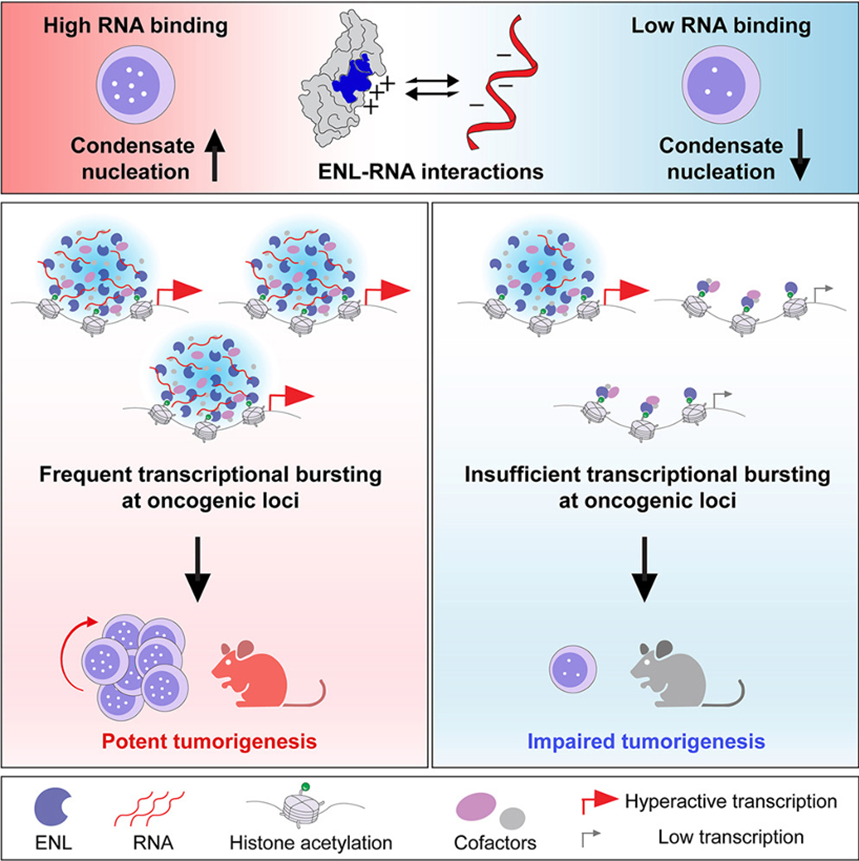
基因表达的精准时空调控,依赖转录因子、表观调控蛋白与染色质微环境的精密协同。近年来,生物分子液 - 液相分离形成的凝聚体成为转录调控领域的研究热点。大量研究证实,细胞核内众多转录调控蛋白可通过相分离形成局部富集的凝聚体,高效招募转录机器至特定基因位点,快速启动基因转录。但当这套调控体系发生病理性紊乱时,异常凝聚体会成为恶性驱动因子,持续放大癌基因表达,助推肿瘤发生发展。
ENL是经典表观组蛋白阅读器,其YEATS结构域特异性识别乙酰化组蛋白,正向调控转录延伸进程。在儿童急性髓系白血病(AML)、肾母细胞瘤中,科研人员已鉴定出一批ENL YEATS结构域功能获得型突变体。李海涛团队联合万里玲团队前期研究已证实,上述突变可驱动ENL在HOXA等致癌靶位点组装异常转录凝聚体,强力激活癌基因转录,介导肿瘤恶性表型。但仍存在一个关键科学问题:除突变本身提升蛋白自聚集能力外,染色质局部微环境是否存在协同因子,辅助致癌凝聚体稳定锚定在Hox这类难转录基因位点,并持续维持高转录活性?
2026 年 6 月 12 日,清华大学基础医学院李海涛团队联合宾夕法尼亚大学医学院万里玲团队,在Molecular Cell期刊发表题为RNA reinforces condensate nucleation on chromatin to amplify oncogenic transcription的研究论文,给出全新调控范式:基因转录产生的新生RNA并非转录过程的“副产物”,而是驱动ENL致癌凝聚体在染色质稳定成核、维持靶基因转录爆发的核心功能性调控分子。该工作系统阐明难转录基因高效转录新机制:新生RNA促进转录凝聚体组装,构建正反馈环路持续强化转录延伸;由此产生的转录放大效应,也为基因延伸阶段的位点特异性调控提供了可靠分子解释。

RNA:从转录旁观者转变为致癌凝聚体重要参与者
RNA在应激颗粒、核仁、核斑点等多种生物凝聚体中承担骨架功能已形成共识,但在转录调控凝聚体体系中,RNA是否参与Hox难转录基因的特异性激活,此前机制尚不清晰。研究团队结合AlphaFold3结构预测、体外EMSA互作实验、细胞CLAP-seq全基因组RNA捕获等手段,证实ENL具备直接RNA结合活性,并精准锁定YEATS结构域一段保守正电荷富集区(K61/R63/K67)为RNA结合核心界面。将该区域三个关键赖氨酸、精氨酸残基突变为丙氨酸构建3A突变体后,ENL与RNA的结合能力大幅衰减,但不破坏其识别乙酰化组蛋白的功能,也不改变蛋白丰度与细胞核定位,为解析RNA介导的表观许可及基因激活机制提供了特异性极强的RNA结合缺陷工具蛋白。
RNA介导凝聚体成核:小分子可逆驱散-重建体系直观验证调控功能
为直观解析RNA对凝聚体成核的调控作用,团队搭建了一套可逆小分子诱导“驱散—重建”实验体系:利用ENL YEATS特异性抑制剂TDI-11055将染色质上已形成的异常凝聚体解离洗脱,撤除药物后实时动态追踪HOXA等位点的凝聚体重建过程。实验结果显示,致癌突变体ENL-T1可在药物洗脱后快速于HOXA位点重建凝聚体,恢复靶基因高水平转录;而RNA结合缺陷3A突变体、或经转录抑制剂阻断新生RNA生成后,凝聚体重成核效率显著下降,Hox等基因的特异性激活水平同步大幅衰减。该体系直接证明:原位新生RNA是凝聚体在靶位点高效成核的关键分子催化剂,也是解锁基因持续高效延伸必不可少的表观许可条件。
全基因组水平证实:RNA的转录放大作用具备严格位点特异性
为进一步探究RNA对ENL突变体的激活效应是全局非特异调控,还是靶向特异性调控,研究人员通过全基因组ChIP-seq图谱解析ENL-T1在染色质的结合分布,发现其结合峰呈极端不均衡分布:仅不足1%的染色质区段呈现超高蛋白富集信号,HOXA簇、CBX3等经典沉默难转录基因均富集于此,研究人员将此类高结合位点定义为 “凝聚体许可位点(top-bound)”,其余位点归为低结合位点(low-bound)。令人意外的是,破坏ENL-RNA互作的3A突变仅选择性削弱top-bound位点的ENL富集与基因转录激活,对low-bound位点几乎无影响;该特征在HEK293、MDA-MB-231两种完全不同的细胞系中均可稳定复现,且不同细胞的凝聚体许可位点基因集存在细胞特异性。上述结果清晰表明,新生RNA的转录促进作用具有精准靶向性,仅定向作用于Hox这类难转录致癌位点,通过转录放大马太效应实现局部转录爆发,而非无差别广谱激活,完美阐释了转录延伸阶段的位点特异性调控逻辑。
造血移植小鼠模型:ENL-RNA互作是白血病恶性转化的核心分子
研究人员在造血干/祖细胞(HSPC)中分别表达野生型ENL-WT、致癌突变ENL-T1、RNA结合缺陷型ENL-T1-3A,开展骨髓移植体内功能验证,体内表型差异极具说服力:ENL-T1-3A虽仍可少量组装凝聚体、激活普通基因,却彻底丧失强效激活HOXA等致癌基因的能力,几乎完全失去致白血病潜能。两组独立移植队列表型高度一致:表达ENL-T1的小鼠全部罹患急性髓系白血病,本质是RNA介导的凝聚体正反馈持续放大Hox癌基因转录,转录马太效应不断累积突破细胞恶性转化阈值;而ENL-T1-3A移植小鼠长达一年观察周期内无一例白血病发病,生存曲线与野生型对照组完全重合。体内实验明确核心结论:ENL突变驱动肿瘤发生存在严格的致癌转录阈值,RNA介导的表观许可通路、凝聚体转录正反馈,是推动Hox难转录基因表达突破阈值的关键环节;仅部分阻断ENL与新生RNA的相互作用,即可打破转录放大马太效应,将癌基因表达压制在致病阈值之下,从根源阻断白血病发生。
总之,该论文提出三层递进式协同致病调控模型,完整解答Hox等难转录基因如何实现特异性活化调控,充分凸显表观许可机制的核心地位:
1. 第一层(内在驱动力):T1-T8等肿瘤相关突变增强ENL分子自聚集能力,赋予蛋白自发形成凝聚体的固有属性;
2. 第二层(染色质锚定):乙酰化组蛋白介导ENL锚定至活跃染色质区域,提供空间定位基础;
3. 第三层(本研究全新发现,表观许可核心环节):原位新生RNA作为关键协同因子,通过与ENL YEATS结构域正电荷区域结合引入多价分子作用力,协同驱动凝聚体在HOXA等难转录靶位点高效成核,搭建RNA -凝聚体-转录延伸正反馈环路,持续驱动靶基因转录爆发,最终诱发白血病等恶性肿瘤。

宾夕法尼亚大学医学院博士生Krista A. Budinich、清华大学基础医学院已毕业博士生姚欣仪和宾夕法尼亚大学医学院博士生龚楚杰为本文共同第一作者。清华大学生命科学学院2026级博士生汪筱婷为本文提供重要帮助。宾夕法尼亚大学医学院万里玲博士、清华大学基础医学院/北京生物结构前沿中心/清华-北大生命联合中心李海涛教授和李元元副研究员为共同通讯作者。该研究工作得到科技部国家重点研发计划和国家自然科学基金委等项目的大力支持。
文章链接
https://doi.org/10.1016/j.molcel.2026.05.024












 科研动态
科研动态